Significación funcional y modelos explicativos
Un elevado
nivel de sincronización entre la actividad cerebral
generada en diferentes estructuras corticales apunta, con una
alta probabilidad, a la existencia de una
relación funcional entre tales regiones. Por lo
tanto, a partir de los datos obtenidos con la técnica
de la coherencia podrían hacerse inferencias acerca de la
distribución
topográfica
de las conexiones córticocorticales implicadas
en la generación, control y/o mantenimiento de un estado cerebral
específico, tanto en sujetos sanos como en pacientes con
determinados cuadros clínicos.
La
posibilidad que ofrece la técnica de la coherencia de cuantificar
cuantificar el grado de asociación
funcional entre áreas cerebrales resulta de
especial interés a nivel clínico, sobre todo si se tiene en
cuenta que tales relaciones tienen su sustrato fisiológico en las
conexiones anatómicas entre redes neuronales localizadas en
diferentes áreas corticales, cuya activación
podría verse seriamente alterada en determinadas
enfermedades cerebrales. En la corteza cerebral
de sujetos humanos pueden identificarse dos sistemas de
fibras de conexión:
1. El que
determina las interacciones a corta distancia, en
el rango del milímetro, debido a la acción de las
interneuronas neocorticales, y 2. Los largos axones que forman
tractos fibrosos encargados de conectar entre sí a grupos
neuronales pertenecientes a regiones corticales
distantes.
Thatcher et
al estudiaron la funcionalidad de ambos sistemas
de fibras corticales en sujetos humanos y establecieron un
modelo de funcionamiento cerebral basado en datos de coherencia
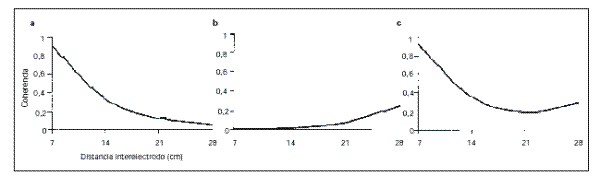
EEG. De acuerdo con este modelo, y según se muestra en la
figura 1, el decremento exponencial de la coherencia a medida
que aumenta la distancia interelectrodo reflejaría la progresiva
desaparición de las conexiones cortas a nivel local
(especialmente
a partir de
14 cm) (Fig. 1a), mientras que el incremento exponencial
de los valores de coherencia entre regiones distantes (mayores
de 14 cm) demostraría la activación de las fibras largas de
conexión córticocortical
entre dichas áreas corticales (Fig. 1b). El
resultado de la acción simultánea de ambos sistemas de fibras de
conexión cortical (cortas y largas) queda evidenciado, por lo
tanto, en la función cuadrática que describen los
datos de coherencia con la distancia, cuya
pendiente negativa inicial ilustraría la influencia
del sistema de conexiones cortas y la pendiente positiva
final, la influencia del sistema de fibras largas (Fig. 1c).
Por otra
parte, en un reciente estudio se ha propuesto que
los datos de coherencia no sólo son sensibles a la existencia de
conexiones anatómicas entre dos regiones cerebrales determinadas,
sino que también podrían proporcionar información sobre las
propiedades espectrales de dichas conexiones. Esto se debe,
fundamentalmente, a que cada red neuronal se
caracteriza por una configuración de subgrupos
neuronales diferente, cada uno de los cuales es
sensible a variar hacia un determinado intervalo de frecuencia.
Así, los
acoplamientos funcionales entre dichas redes, a
través de las fibras anatómicas de conexión, se establecen cuando
en ellas existen subgrupos neuronales que oscilan a la misma
frecuencia. Por lo tanto, el espectro de coherencia entre dos
regiones corticales reflejará para qué grupos de
osciladores de una red neuronal se establecen
conexiones funcionales en un determinado estado
de activación cerebral. Un claro ejemplo de la ventaja que
supone el adoptar esta perspectiva se encuentra en el estudio
realizado por Achermann y Borbély, donde, gracias
a la utilización de una resolución espectral
importante (0,25 Hz) y al uso de
filtros con
muy baja frecuencia de corte (0,16 Hz), se detectó una
significativa sincronización de los componentes más lentos de la
banda delta (0,751,5 Hz) y de la actividad
de husos de sueño (1214 Hz)
sobre amplias regiones de la corteza durante el sueño
no REM de sujetos adultos sanos, hecho que parece reflejar la
acción de dos osciladores diferentes a nivel talamocortical.
Además del
aumento de la resolución espectral asociada a la
observación simultánea de todo el espectro de coherencia obtenido
a partir de dos derivaciones cualesquiera, existen otras formas
de optimizar la utilidad de esta técnica de análisis. La
composición espectral o el estudio de la
conectividad córticocortical durante
el sueño se ha realizado tradicionalmente atendiendo a cada una
de sus fases. No obstante, este enfoque conlleva un importante
coste relacionado con la pérdida de información contenida en los
numerosos microestados cerebrales que se suceden a lo
largo de una misma fase del sueño. Así, se ha demostrado, por
ejemplo, que la coherencia interhemisférica durante la fase REM es
mayor en períodos de actividad oculomotora que en períodos
tónicos. El
hecho de que este incremento se observe incluso sobre
electrodos posteriores invita a descartar la posibilidad de que se
deba a contaminación artefactual causada por la actividad oculomotora típica
de la fase REM. En su lugar, se ha propuesto que este
resultado es un reflejo del mayor nivel de activación de las fibras
callosas durante períodos de fase REM con movimientos oculares rápidos en
comparación con períodos de quietud oculomotora.
Por otra parte, una misma actividad cerebral puede
mostrar un patrón de coherencia diferente en función del estado
cerebral donde esta se registre. En este sentido, Cantero et al
demostraron que el ritmo alfa que aparece en una vigilia relajada
con ojos cerrados, la actividad alfa de somnolencia y los brotes
de alfa que surgen espontáneamente durante la fase REM, muestran un
nivel de sincronización diferente entre regiones fronto-occipital
y frontofrontales a medida que va disminuyendo el nivel de activación
cerebral; así se presentan valores mayores de coherencia
en vigilia, menores en somnolencia y mínimos en la fase REM.
Otro aspecto que, sin duda, va a determinar la validez
de las interpretaciones funcionales de los datos de coherencia
está relacionado con la variabilidad
intraindividual. Partiendo del hecho de que las características del EEG son altamente
variables entre sujetos, recurrir a muestras amplias facilitará
que los valores de coherencia sean más representativos del patrón de
relaciones funcionales característico de un estado cerebral
determinado. En lo referente a variaciones intraindividuales, existen
estudios que muestran una elevada estabilidad de los valores de
coherencia obtenidos para un único sujeto a lo largo de sucesivas
sesiones experimentales.
No obstante, en uno de estos estudios
pudo comprobarse, además, que el grado de
consistencia de los resultados de las 11 sesiones de vigilia relajada
realizadas a lo largo de un mes dependía de la banda de frecuencia
considerada.
Concretamente, la banda alfa fue la que mostró un
comportamiento más inestable a lo largo del tiempo, lo cual sugería una
mayor sensibilidad de esta actividad cerebral a factores
potencialmente ajenos a la condición que se pretende estudiar, como por
ejemplo, la presencia/ausencia de una determinada enfermedad.
Por último, además de tener en cuenta todos los factores
anteriormente comentados, es necesario asumir algunas limitaciones
limitaciones a la hora de interpretar los acoplamientos funcionales
entre áreas corticales a partir de los datos obtenidos con la
técnica de la coherencia. Entre ellas destacan los inconvenientes
derivados de la técnica del EEG mediante la utilización de electrodos
de superficie.
En este sentido, podría ocurrir que, a pesar de que dos
fuentes generadoras de una misma actividad cerebral se
relacionaran funcionalmente, la coherencia entre las señales EEG
registradas sobre las localizaciones correspondientes a cada uno de
estos generadores fuera muy baja, simplemente por el hecho de
que uno
de ellos tuviera una orientación tangencial con respecto
a la superficie cortical. En este caso, el uso complementario de
técnicas de registro neuromagnéticas resolverían el problema, ya que
ofrecen una excelente resolución espacial y temporal de la
actividad cerebral generada por dipolos tangenciales. Igualmente, es
necesario tener en cuenta que la actividad generada a nivel
subcortical alcanza amplias áreas de la superficie cortical, bien por su
expansión pasiva, bien por la existencia de vías de conexión
talamocortical, lo que provocaría la obtención de altos valores de
coherencia entre regiones distantes que serían erróneamente interpretados
de acuerdo con acoplamientos funcionales entre estructuras
corticales.
Una vez expuestas algunas de las cuestiones más
importantes relacionadas con la interpretación funcional de los
datos de coherencia, se expondrán a continuación algunos de los objetivos
experimentales y clínicos que la aplicación de la técnica de la
coherencia ha permitido cubrir. En primer lugar, y dado que esta
técnica es sensible al grado en que las interacciones
neocorticales a corta y larga distancia contribuyen a la génesis de una
actividad determinada, su uso permitiría estudiar el
patrón topográfico de interacciones córticocorticales
subyacente a la generación de una misma actividad en diferentes estados de
activación cerebral (Cantero et al, datos no publicados).
En segundo lugar, dicha técnica puede emplearse en la evaluación de
enfermedades que conlleven una degeneración cortical progresiva, como
ocurre en las demencias, o la pérdida de determinados
tractos fibrosos, como en los especialmente llamativos
casos de pacientes con ausencia del cuerpo calloso. Un
tercer campo de aplicación sería la evaluación de los cuadros
clínicos asociados con alteraciones del funcionamiento normal de
diferentes estructuras cerebrales que generan o controlan
determinados estados fisiológicos y que, como consecuencia, conllevan
irregularidades en el establecimiento de conexiones entre estas y otras
estructuras. No obstante, antes de
adentrarnos en los estudios que han mostrado el valor clínico
de la técnica de la coherencia EEG durante los diferentes estados cerebrales que componen
el sueño, revisaremos los trabajos que se refieren al patrón de coherencia durante
el sueño en sujetos humanos sanos.

 Enlaces
Seleccionados
Enlaces
Seleccionados